[강석기의 과학카페] 마침내 밝혀진 냄새수용체 구조

1990년 인간게놈프로젝트가 시작됐을 때만 해도 단백질 지정 유전자 개수가 10만 개는 될 것이라고 봤다. 그런데 작업이 진행되면서 예상값이 점점 줄어들었고 해독 결과 불과 2만 개라는 충격적인 사실이 밝혀졌다. 이에 대해 중요한 건 유전자 개수가 아니라 유전자 사이의 네트워크의 복잡성이라고 설명한다.
실제 다른 유전자의 발현을 조절하는 전사인자를 지정하는 유전자가 2000여 개로 전체의 10%를 넘는다. 세포 안팎의 신호를 전달하는 데 관여하는 유전자도 그 정도 되는데, 호르몬 같은 신호분자(리간드라고 부른다)의 유전자와 이를 인식하는 수용체 유전자로 이뤄져 있다.
한편 수용체의 대다수가 G단백질연결수용체(GPCR)로 거의 1000가지나 된다. GPCR은 세포막에 박혀있는 단백질로 바깥쪽에서 온 리간드가 달라붙으면 구조가 바뀌면서 세포 안쪽의 G단백질에 영향을 줘 신호를 전달한다. 시각, 후각, 미각 같은 감각 신호(각각 광자(빛)와 냄새분자, 맛분자)도 GPCR을 통해 전달된다. 특히 냄새수용체 유전자는 거의 400가지나 돼 전체 유전자의 2% 가까이 차지하고 있다.
● 냄새분자 붙은 다른 수용체 구조는 밝혔지만
신호 전달에 문제가 있는 질병에서 GPCR은 약물의 주요 표적이라는 점에서 구조를 밝히는 연구가 치열했다. 2000년 빛을 감지하는 GPCR인 로돕신의 구조가 처음 밝혀졌고 2007년 스트레스 호르몬인 아드레날린의 신호를 전달하는 베타-아드레날린성 수용체의 구조가 규명됐다. 그 뒤 80가지가 넘는 GPCR의 구조가 밝혀졌다. 그런데 뜻밖에도 GPCR의 거의 절반을 차지하는 냄새수용체의 구조는 하나도 밝혀진 게 없다. 신약 개발 같은 동기부여가 약하다는 걸 감안해도 너무한 게 아닐까.

흥미롭게도 냄새수용체의 리간드, 냄새분자가 다른 수용체에 달라붙어 있는 상태의 구조는 밝혀졌다. 박하향을 내는 분자인 멘톨은 냄새수용체인 OR8K3에 달라붙어 달콤한 향기를 느끼게 하지만 동시에 냉각수용체인 TRPM8에도 결합해 시원하다는 느낌을 준다. 원래 TRPM8은 온도가 낮아지면 구조가 바뀌면서 활성화돼 신호가 전달되는 것인데, 멘솔 분자가 달라붙으면 상온에서도 활성화돼 신호를 보내 뇌가 ‘온도가 낮다’고 착각하게 만든다.
지난 2020년 TRPM8의 비활성 상태 구조와 멘톨이 달라붙어 활성화된 구조를 밝힌 연구 결과가 나왔다. 그럼에도 정작 멘톨의 진짜 파트너인 냄새수용체 OR8K3의 구조는 아직 나오지 않았다. 리간드가 알려진 다른 냄새수용체도 마찬가지인데, 냄새수용체 분자는 구조의 변동 폭이 커 구조 규명 연구에 어려움이 크다고 한다.
● 식초의 시큼한 냄새가 느껴질 때 일어나는 일
지난 15일 학술지 ‘네이처’는 처음으로 냄새수용체 구조를 밝힌 논문을 사이트에 미리 공개했다. 샌프란시스코 캘리포니아대 등 미국의 공동연구자들은 식초의 주성분인 아세트산의 시큼한 냄새를 매개하는 냄새수용체 OR51E2에 프로피온산이 달라붙어 활성화된 상태의 구조를 밝히는 데 성공했다. 프로피온산은 아세트산보다 탄소원자 하나가 더 많은 분자로 치즈의 시큼한 냄새를 내는 주인공이다. 둘 다 같은 냄새수용체(OR51E2)에 작용한 결과다.
GPCR은 아미노산 사슬이 나선을 그리며 일곱 차례에 걸쳐 세포막을 관통하며 박혀있는 단백질이다. 바느질로 비유하면 천 바깥에서 바늘을 찔러 통과시키고 다시 안에서 밖으로 바늘을 찔러 통과시키는 식으로 일곱 번을 반복해 실이 천에 고정된 셈이다.
지금까지 구조가 밝혀진 GPCR로 알아낸 작동 메커니즘은 이렇다. 막을 관통해 박혀있는 일곱 개의 나선 사이에 생긴 공간에 리간드가 들어가 자리를 잡으면 나선들이 재배치되면서 구조가 살짝 바뀌어 세포막 안쪽에 있는 G단백질을 활성화해 신호가 전달된다.
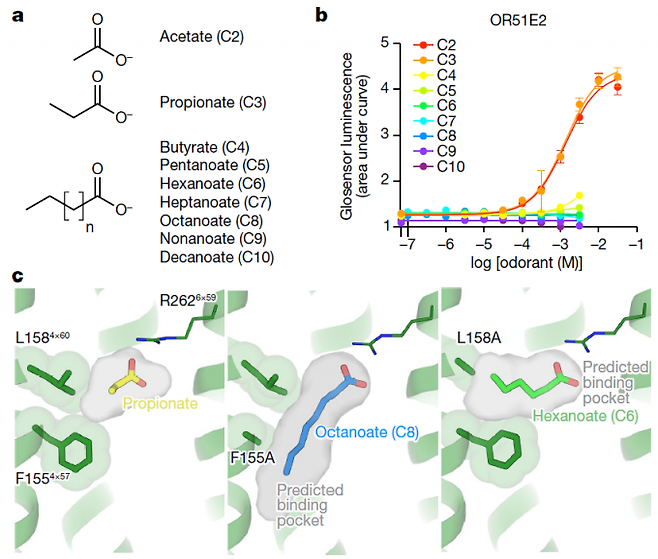
연구자들은 극저온전자현미경분석법으로 OR51E2의 3차원(입체) 구조를 분석했다. 리간드인 프로피온산 없이 수용체 단백질만 있는 비활성 상태의 구조는 아쉽게도 얻지 못했다. 구조의 유동성이 워낙 커서 “이거다!”라고 하나를 정할 수가 없었기 때문이다.
리간드와 G단백질이 결합해 활성화된 상태인 OP51E2의 구조는 변동 폭이 작아 밝혀낼 수 있었다. 분석 결과 프로피온산 분자는 OP51E2의 막관통 나선 3, 4, 5, 6번 사이의 공간에 쏙 들어가 있었다. 이 나선들에 있는 몇몇 아미노산이 프로피온산과 상호작용해 안정화시킨 결과다.
공간의 크기는 탄소원자 2개인 아세트산과 3개인 프로피온산은 들어갈 수 있지만 4개인 뷰티르산부터는 들어가지 못하고 따라서 이들 큰 분자는 OP51E2를 활성화하지 못한다. 흥미롭게도 아미노산을 바꿔치기해 공간을 넓혀주자 아세트산과 프로피온산 같은 작은 분자는 너무 헐거워 붙지 못하고 탄소원자 8개인 카프릴산 분자가 오히려 꼭 맞아 수용체를 활성화했다.
그렇다면 비활성 상태인 수용체에 리간드가 붙어 활성화될 때 구조가 어떻게 바뀌는 걸까. 아쉽게도 비활성 상태 구조를 분석하는 데는 실패했지만 대신 베타-아드레날린성 수용체의 비활성 구조를 참고해 컴퓨터시뮬레이션으로 구조를 예측했다. 아울러 요즘 한창 인기인 단백질 구조 예측 AI 프로그램인 알파폴드2도 활용했다. 담난 알파폴도2 역시 냄새수용체 구조 연구에서는 정확도가 떨어져 보조적 역할에 머물렀다.
그 결과 얻은 비활성 상태 구조와 앞서 현미경으로 규명한 활성 상태 구조를 비교한 결과 리간드가 결합했을 때 변화를 제안하는 모델을 만들 수 있었다. 흥미롭게도 세포막 바깥쪽에 놓인, 막관통 나선 6번과 7번 사이를 연결하는 부분이 수용체가 활성화되는 데 중요한 역할을 하는 것으로 밝혀졌다.

● 냄새분자 사이 작용 복잡해
이번 연구로 냄새수용체 작동 미스터리가 풀리기 시작했지만 아직 갈 길은 멀다. 우리가 냄새를 지각하는 방식이 꽤 복잡하기 때문이다. 리간드(냄새 분자)와 냄새수용체가 1:1로 대응하는 게 아니라(그렇다면 냄새 분자는 기껏해야 400개에 불과할 것이다) 한 리간드가 여러 수용체와 결합하고 한 수용체에도 여러 리간드가 붙을 수 있기 때문이다.

대부분의 냄새는 여러 냄새분자가 여러 수용체와 복잡하게 작용한 결과다. 얼핏 생각하면 이 과정은 합집합의 법칙을 따를 것 같다. 예를 들어 냄새분자 A가 냄새수용체 10가지를 활성화하고 분자 B가 수용체 15가지를 활성화하고 둘이 겹치는 수용체가 5개라면(교집합), A와 B 혼합물의 냄새는 수용체 20개가 활성화된 결과다(10+15-5=20).
이런 식이면 냄새분자를 더할수록 점점 더 많은 수용체가 활성화돼 결국은 모든 수용체가 활성화됐을 때 느끼는 냄새(그런 게 있다면)에 가까워질 것이다. 그런데 실제로는 이런 일이 일어나지 않는다. 냄새분자는 어떤 냄새수용체는 활성화시키지만 다른 냄새수용체는 오히려 활성을 억제하기 때문이다. 향수를 만들 때 여러 향료를 섞어도 오히려 향이 부드러워지는 현상도 이 때문이다.
척추동물의 냄새수용체는 크게 두 가지로 나뉘는데, 수억 년 전 바다에만 살던 시절 진화한 1군과 육상으로 올라온 뒤 진화한 2군이다. 1군은 물(바다)에 녹아 있는 냄새 분자를, 2군은 공기 중에 떠다니는 냄새 분자를 감지하게 진화했다는 말이다. 사람의 냄새수용체는 1군이 약 60가지로 15%에 불과하고 나머지 85%는 2군이다.
이번에 구조가 밝혀진 OR51E2는 물에 잘 녹는 아세트산과 프로피온산을 감지하는 1군 냄새수용체다. 꽃향기를 비롯해 냄새의 다채로운 스펙트럼을 담당하는 2군 냄새수용체는 구조의 불안정성이 더 큰 것으로 알려져 있다.
추가적인 구조 연구를 통해 냄새분자가 어떻게 특정 냄새수용체에서는 활성화를 촉진하고 다른 수용체에서는 억제하는가를 알아내야 후각에 대한 진정한 이해에 도달할 것이다. 앞으로도 갈 길이 멀다는 얘기다.
※ 필자소개
강석기 과학칼럼니스트 (kangsukki@gmail.com). LG생활건강연구소에서 연구원으로 근무했고 2000년부터 2012년까지 동아사이언스에서 기자로 일했다. 2012년 9월부터 프리랜서 작가로 활동하고 있다. 직접 쓴 책으로 《강석기의 과학카페》(1~7권),《생명과학의 기원을 찾아서》가 있다. 번역서로는 《반물질》, 《가슴이야기》, 《프루프: 술의 과학》을 썼다.
[강석기 과학 칼럼니스트 kangsukki@gmail.com]
Copyright © 동아사이언스. 무단전재 및 재배포 금지.